Introducción
La verdad es que la biofluorescencia es una característica que poseen un gran número de vertebrados, los cuales tienen esa curiosa capacidad de iluminarse con ciertos tipos de luz. Pero nunca antes me había planteado la siguiente pregunta: si algunas aves son fluorescentes y sabemos que las aves son dinosaurios. ¿Quiere decir esto que los dinosaurios no avianos también fueron fueron fluorescentes?
 |
| Un frailecillo atlántico (Fratercula arctica) con su pico fluorescente. Créditos de la imagen: Jamie Dunning |
¿Qué significa ser fluorescente y bioluminiscente?
Desde principios de la década de 1980, se sabe que algunas aves pueden detectar longitudes de onda UV (Bennett y Cuthill, 1994; Bowmaker et al., 1997; Wikie et al., 1998; Cuthill et al., 2000) y estudios más recientes han demostrado que se trata de una característica bastante común entre los múltiples linajes de vertebrados, como peces (Sparks et al., 2014), reptiles (Font y Molina-Borja, 2004; Protzel et al., 2018), anfibios (Cox y Fitzpatrick, 2023; Lamb y Davis, 2020; Thompson et al., 2020) e incluso en mamíferos (Kohler et al., 2019). Y, cómo no olvidarnos de nuestros queridos invertebrados (Lim et al., 2007).La biofluorescencia es un fenómeno que permite que ciertos organismos, absorban luz en una longitud de onda y que luego la emita en una longitud de onda más larga, lo que hace que brillen en colores visibles.
Mientras que la bioluminiscencia es una reacción química mediante la cual un organismo genera su propia luz, como ocurre en las luciérnagas (Marques y Esteves da Silva, 2009) o ciertos peces abisales que llevan a cabo una relación simbiótica con bacterias para producir luz (Haygood, 1993)
Mientras que la bioluminiscencia es una reacción química mediante la cual un organismo genera su propia luz, como ocurre en las luciérnagas (Marques y Esteves da Silva, 2009) o ciertos peces abisales que llevan a cabo una relación simbiótica con bacterias para producir luz (Haygood, 1993)
La revolución de la fluorescencia
Pero centrémonos en las aves. Muchas especies poseen plumaje que refleja longitudes de onda UV. Como los pinguinos (Thomas et al., 2013), los búhos nocturnos y los chotacabras (Weidensaul et al., 2011; Camacho et al., 2019), los loros (Hausman et al., 2003) y las avutardas (Galvan et al., 2016).
Aunque las plumas no son la única parte del cuerpo que puede exhibir fluorescencia. Se ha documentado en otras partes del cuerpo en el mérgulo crestado (Aethia cristatella) (Wails et al., 2017), el frailecillo atlántico (Fratercula arctica) (Dunning et al., 2018) y el mérgulo rinoceronte (Cerorhinca monocerata) (Wilkinson et al., 2019).
 |
| Aves que presentan florescencia en sus picos. De izquierda a derecha: mérgulo crestado (Aethia cristatella), frailecillo atlántico (Fratercula arctica) y mérgulo rinoceronte (Cerorhinca monocerata). Créditos de las imágenes en el mismo orden: James A. Giroux; Sophie Roy y Kyle Bowman. |
Pero, ¿por qué las aves desarrollarían esta característica? Se ha planteado que la fluorescencia en el plumaje, así como en otra partes del cuerpo, puede cumplir una función en la señalización social y sexual, siendo únicamente el grupo de los loros, alcas y frailecillos los únicos que han sugerido un rol potencial de la biofluorescencia como señal visual en el cortejo y la cópula.
Algunas especies de loros (Psittaciiformes), tienen un plumaje fluorescente que absorbe longitudes de onda cortas (UV o azul) y las reemite en longitudes de onda más largas, haciendo que el plumaje "brille" (Boles, 1990; Pearn et al., 2001; Arnold et al., 2002). Por ejemplo, el periquito australiano (Melopsittacus undulatus) puede usar el plumaje biofluorescente en las exhibiciones de cortejo como señal o para mejorar el contraste contra otro plumaje altamente visible que refleja la luz UV (Pearn et al., 2001; Arnold et al., 2002)
 |
| Periquito australiano (Melopsittacus undulatus). Créditos de la imagen: Ciencia Verde |
Curiosamente, también se ha observado una correlación positiva entre las plumas fluorescentes y los plumajes utilizados en las exhibiciones de cortejo, los cuales a menudo se acompañan de un plumaje que refleja los rayos UV (Hausmann et al., 2003).
Mientras que en el caso de las alcas y los frailecillos, la biofluorescencia se produce en el cere, el pico y placas del pico. Incluso, se ha descubierto que el frailecillo cornudo (Fratercula corniculata) presenta un dimorfismo sexual en el tamaño de los parches biofluorescentes (Evtukh, 2019)
Se cree que las estructuras visibles en estos grupos tienen una función de señalización reproductiva, y las emisiones biofluorescente adicionales pueden servir como una señal visual.
Asimismo, recientemente se ha descubierto la presencia de plumas fluorescentes, así como otras partes de su cuerpo en las aves del paraíso (Paradisaeidae). En los machos, la biofluorescencia se presenta en el plumaje brillante y la piel, que se destacan y utilizan en las exhibiciones de arena y cortejo (Martin et al., 2025). Las ubicaciones de estas áreas biofluorescentes varían en función de la especie, pero incluye la boca interna, el pico, las plumas de la cabeza, el cuello, las patas y los penachos.
En la gran mayoría de los casos, ya sea por la presencia de un plumaje constrastante o por el comportamiento del individuo, las áreas biofluorescentes del cuerpo contrastan marcadamente y destacan en el ambiente.
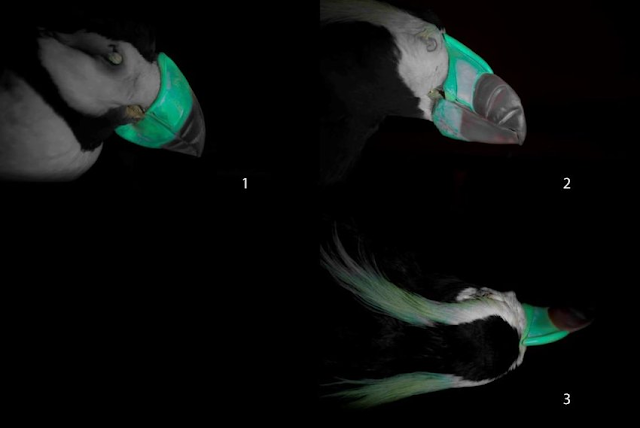 |
| Área de brillo fluorescente en: 1-Pico de frailecillo cornudo; 2-Pico de frailecillo copetudo; 3-Tobas de frailecillo copetudo. De Evtukh, 2019. |
Se cree que las estructuras visibles en estos grupos tienen una función de señalización reproductiva, y las emisiones biofluorescente adicionales pueden servir como una señal visual.
Asimismo, recientemente se ha descubierto la presencia de plumas fluorescentes, así como otras partes de su cuerpo en las aves del paraíso (Paradisaeidae). En los machos, la biofluorescencia se presenta en el plumaje brillante y la piel, que se destacan y utilizan en las exhibiciones de arena y cortejo (Martin et al., 2025). Las ubicaciones de estas áreas biofluorescentes varían en función de la especie, pero incluye la boca interna, el pico, las plumas de la cabeza, el cuello, las patas y los penachos.
 |
| Ejemplo de ochos especies diferentes de ave del paraíso macho fotografiadas bajo luz blanca (arriba) y que también muestran regiones biofluorescentes (abajo). De Martin et al., 2025 |
En la gran mayoría de los casos, ya sea por la presencia de un plumaje constrastante o por el comportamiento del individuo, las áreas biofluorescentes del cuerpo contrastan marcadamente y destacan en el ambiente.
Aunque se desconoce la visión en aves del paraíso, en base a la inferencia filogenética se sugiere que los machos y las hembras probablemente solo son sensibles al violeta e incapaces de visualizar la luz UV (Odeen et al., 2011). Pero los picos espectrales de dos de los conos de sus ojos coinciden con los de sus emisiones biofluorescentes.
No obstante, podríamos plantear que la fluoresencia es un subproducto de otros fenómenos biológicos y no tener que estar relacionado con una función de comunicación o de señalización sexual. Entonces...
En el caso de los búhos, la ausencia de fluorescencia visible en la superficie dorsal, la cabeza o el rostro, lo que indica que es poco probable que se utilice como una señal social (Weidensaul et al., 2011). Aunque las superficie de la parte inferior de las alas sería visibles en vuelo, durante los rituales de cortejo, Pero la falta de conos sensibles al ultravioleta/violeta descarta por completo esta función (Bowmaker y Martin, 1978; Cuthill et al., 2000)
Mientras que en el caso de los pinguinos, se sabe que las manchas del pico de los pinguinos rey (Aptenodytes patagonicus) y emperador (Aptenodytes forsteri) tienen reflectancias en las manchas anaranjadas de sus picos, por lo que se cree que podría estar asociado con la madurez sexual y los comportamientos de cortejo (Jouventin et al., 2005). Esta reflectancia es más alta en los pinguinos de apareamiento temprano, lo que podría estar relacionado con el éxito reproductivo (Jouventin et al., 2009; Nolan et al., 2010).
Pero, a pesar de encontrar reflectancias de UV en el pico de estas dos especies, la reducción en la transmisión de luz UV en los ojos de los pinguino rey, hace que sea poco probable que desempeñe un papel importante en la selección de pareja (Hadden et al., 2023).
No obstante, podríamos plantear que la fluoresencia es un subproducto de otros fenómenos biológicos y no tener que estar relacionado con una función de comunicación o de señalización sexual. Entonces...
¿Qué nos indica que la fluorescencia puede tener una función de comunicación?
Marshal y Johnsen, 2017 y Nicolai et al., 2024, sugieren utilizar una serie de criterios para poder evaluar si la biofluorescencia en un organismo tienen una función visual significativa para los individuos que la exhiben: (I) Las señales fluorescentes deben ser visibles para otros individuos de la misma especie o incluso de diferentes especies. (II) Las emisiones fluorescentes deben destacar en su entorno natural. (III) Las zonas del cuerpo donde se presentan la fluorescencia deben de estar asociadas a comportamientos de señalización, como la atracción de pareja o la defensa del territorio. (VI) Los receptores visuales de los animales deben de ser sensibles a las longitudes de onda específica de la luz fluorescente emitida. Por estas razones, otros grupos de aves, como los búhos y los pinguinos, a pesar de que presentan fluorescencia, no cumplen con los criterios establecidos.
En el caso de los búhos, la ausencia de fluorescencia visible en la superficie dorsal, la cabeza o el rostro, lo que indica que es poco probable que se utilice como una señal social (Weidensaul et al., 2011). Aunque las superficie de la parte inferior de las alas sería visibles en vuelo, durante los rituales de cortejo, Pero la falta de conos sensibles al ultravioleta/violeta descarta por completo esta función (Bowmaker y Martin, 1978; Cuthill et al., 2000)
 |
| Vistas emparejadas de las superficies ventrales de las alas de un búho. Fotografías de Scott Weidensaul. De Weidensaul et al., 2011). |
Mientras que en el caso de los pinguinos, se sabe que las manchas del pico de los pinguinos rey (Aptenodytes patagonicus) y emperador (Aptenodytes forsteri) tienen reflectancias en las manchas anaranjadas de sus picos, por lo que se cree que podría estar asociado con la madurez sexual y los comportamientos de cortejo (Jouventin et al., 2005). Esta reflectancia es más alta en los pinguinos de apareamiento temprano, lo que podría estar relacionado con el éxito reproductivo (Jouventin et al., 2009; Nolan et al., 2010).
Pero, a pesar de encontrar reflectancias de UV en el pico de estas dos especies, la reducción en la transmisión de luz UV en los ojos de los pinguino rey, hace que sea poco probable que desempeñe un papel importante en la selección de pareja (Hadden et al., 2023).
¿Es posible la fluorescencia en dinosaurios no avianos?
La presencia de órbitas grandes y lóbulos ópticos bien desarrollado demuestran que los dinosaurios no avianos y los pterosaurios tenían ojos grandes y buena visión. Pudiéndose suponer que tenían un estilo de vida crepuscular (Longrich, 2010). Sin embargo, el tamaño relativo de los ojos ni la agudeza visual demuestra la capacidad de percibir colores, por lo que nuestro único recurso es el encuadre filogenético.
El encuadra filogenético es un método que permite deducir características de organismos extintos a partir del estudio de sus parientes vivos más cercanos (Witmer y Thomason, 1995). En este caso, los arcosaurios actuales, que incluyen a los cocodrilos y aves, sirven como referencia. Mientras que los cocodrilos son tricromáticos, la mayoría de las aves son tetracromáticas. Desde un punto de vista filogenético, esto podría parecer contradictorio, pero no, algunas tortugas, que son parientes cercanas de los arcosaurios, también poseen visión tetracromática (Twyman et al., 2016).
Además los cocodrilos podrían haber evolucionado secundariamente hacia la tricomacia (Kelber et al., 2003), lo que sugiere que los arcosaurios más basales pudieron haber sido tetracromáticos. Esta idea se ve apoyada por estudios moleculares que sitúan a las tortugas y arcosaurios como grupos hermano dentro del clado Archelosauria (Chiari et al., 2012; Crawford et al., 2015). En este sentido, podemos especular que los arcosaurios extintos probablemente veían en color y que la sensibilidad a la luz ultravioleta, aunque no pueda demostrarse directamente, es probable para estos animales, dada su presencia en sus parientes actuales.
Dado que la visión UV es predominante en los diápsidos, se considera altamente probable que los dinosaurios no avianos fueran tetracromáticos y que algunos utilizaran estructuras queratinosas con propiedades biofluorescentes en exhibiciones visuales. Sin embargo, la función exacta de esta posible biofluorescencia en dinosaurios sigue siendo puramente especulativa.
Ahora bien, en caso de presentar biofluorescencia, ¿qué estructuras podrían haber sido biofluorescentes? Esencialmente, se piensa que todas las estructuras asociadas con la exhibición visual en pterosaurios y dinosaurios no aviares estaban recubiertas de queratina. Las crestas craneales son comunes en los pterosaurios (Hone et al., 2011), al igual que las vainas distales y otros apéndices caudales, los cuales podrían haber desempañado algún papel en la comunicación.
También otras estructuras que podrían haber sido estructuras de exhibición son los cuernos, las protuberancias, domos, crestas, placas, cascos, volantes y velas dorsales, son generalmente considerados como estructuras de exhibición visual, aunque otras funciones como la disipación del calor podrían haber jugado roles secundarios.
Sauropodomorpha se considera el único clado dinosauriano que no desarrolló ninguna forma de estructura de exhibición, pero las espinas dérmicas de algunos diplodocoides, las estructuras internariales elevadas de algunos macronarios (Hone et al., 2011), así como los osteodermos cubiertos de queratina de algunos titanosaurios podrían haber funcionado en estos roles de exhibición.
Los ornitischios de algunas líneas poseían cerdas o estructuras más complejas y ramificadas, parecidas a plumas, como Kulindadromeus (Godefroit et al., 2014). Los pterosaurios también poseían filamentos integumentarios, que en algunas líneas, son ramificados y similares a plumas (Yang et al., 2019). Cualquier o todas estas estructuras integumentarias podrían haber funcionado en la exhibición visual y, por lo tanto, dependiendo de las capacidades visuales del animal, podrían haber tenido propiedades biofluorescentes
El encuadra filogenético es un método que permite deducir características de organismos extintos a partir del estudio de sus parientes vivos más cercanos (Witmer y Thomason, 1995). En este caso, los arcosaurios actuales, que incluyen a los cocodrilos y aves, sirven como referencia. Mientras que los cocodrilos son tricromáticos, la mayoría de las aves son tetracromáticas. Desde un punto de vista filogenético, esto podría parecer contradictorio, pero no, algunas tortugas, que son parientes cercanas de los arcosaurios, también poseen visión tetracromática (Twyman et al., 2016).
 |
| Filogenia de Vertebrata con los clados que presentan fololuminiscencia resaltados en azul neón. Elaborado por Ciencia Verde. |
Además los cocodrilos podrían haber evolucionado secundariamente hacia la tricomacia (Kelber et al., 2003), lo que sugiere que los arcosaurios más basales pudieron haber sido tetracromáticos. Esta idea se ve apoyada por estudios moleculares que sitúan a las tortugas y arcosaurios como grupos hermano dentro del clado Archelosauria (Chiari et al., 2012; Crawford et al., 2015). En este sentido, podemos especular que los arcosaurios extintos probablemente veían en color y que la sensibilidad a la luz ultravioleta, aunque no pueda demostrarse directamente, es probable para estos animales, dada su presencia en sus parientes actuales.
Dado que la visión UV es predominante en los diápsidos, se considera altamente probable que los dinosaurios no avianos fueran tetracromáticos y que algunos utilizaran estructuras queratinosas con propiedades biofluorescentes en exhibiciones visuales. Sin embargo, la función exacta de esta posible biofluorescencia en dinosaurios sigue siendo puramente especulativa.
Ahora bien, en caso de presentar biofluorescencia, ¿qué estructuras podrían haber sido biofluorescentes? Esencialmente, se piensa que todas las estructuras asociadas con la exhibición visual en pterosaurios y dinosaurios no aviares estaban recubiertas de queratina. Las crestas craneales son comunes en los pterosaurios (Hone et al., 2011), al igual que las vainas distales y otros apéndices caudales, los cuales podrían haber desempañado algún papel en la comunicación.
 |
| Representación artística de Pteranodon, presentado una cola que podría haber desempeñado algún papel en la comunicación. Créditos de la imagen: Hodarinundu. |
También otras estructuras que podrían haber sido estructuras de exhibición son los cuernos, las protuberancias, domos, crestas, placas, cascos, volantes y velas dorsales, son generalmente considerados como estructuras de exhibición visual, aunque otras funciones como la disipación del calor podrían haber jugado roles secundarios.
Sauropodomorpha se considera el único clado dinosauriano que no desarrolló ninguna forma de estructura de exhibición, pero las espinas dérmicas de algunos diplodocoides, las estructuras internariales elevadas de algunos macronarios (Hone et al., 2011), así como los osteodermos cubiertos de queratina de algunos titanosaurios podrían haber funcionado en estos roles de exhibición.
 |
| Neuquensaurus australis, un titanosaurio que presenta osteodermos. Créditos de la imagen: Carlos sapiens. |
Los ornitischios de algunas líneas poseían cerdas o estructuras más complejas y ramificadas, parecidas a plumas, como Kulindadromeus (Godefroit et al., 2014). Los pterosaurios también poseían filamentos integumentarios, que en algunas líneas, son ramificados y similares a plumas (Yang et al., 2019). Cualquier o todas estas estructuras integumentarias podrían haber funcionado en la exhibición visual y, por lo tanto, dependiendo de las capacidades visuales del animal, podrían haber tenido propiedades biofluorescentes
Conclusión
Y pues esto ha sido todo. Realmente, este artículo no es más que un conjunto de especulaciones y sugerencias, pero basándose en las pruebas filogenéticas, es una idea valida. A veces, la literatura científica no debería de centrarse únicamente en los resultados, sino también en el debate y la especulación, ya que de esta manera se puede obtener "más" información que a partir de los resultados por sí solos. Siempre debe de haber un poco de imaginación, pero aplicada de manera lógica y coherente, en función de las pruebas existentes
Para otras publicaciones sobre biofluorescencia, consulte...
Para otras publicaciones sobre biofluorescencia, consulte...
-Pelaje brillante (Junio, 2025)
Bibliografía
- Marques, Simone & Silva, Joaquim. (2008). Firefly Bioluminescence: A Mechanistic Approach of Luciferase Catalyzed Reactions. IUBMB life. 61. 6-17. 10.1002/iub.134. https://iubmb.onlinelibrary.wiley.com/doi/full/10.1002/iub.134
- Haygood, M. G. (1993). Light a Organ Symbioses in Fishes. Critical Reviews in Microbiology, 19(4), 191–216. https://doi.org/10.3109/10408419309113529
- BENNETT,A.T.AND I. C. CUTHILL. 1994. Ultraviolet visionin birds: what is its function?
- BOWMAKER, J. K., L. A. HEATH,S.E.WILKIE,AND D. M.HUNT. 1997. Visual pigments and oil droplets from sixclasses of photoreceptor in the retinas of birds.
- Sparks, John & Schelly, Robert & Smith, W & Davis, Matthew & Pieribone, Vincent & Gruber, David. (2014). The Covert World of Fish Biofluorescence: A Phylogenetically Widespread and Phenotypically Variable Phenomenon. PloS one. https://www.researchgate.net/publication/259722578_The_Covert_World_of_Fish_Biofluorescence_A_Phylogenetically_Widespread_and_Phenotypically_Variable_Phenomenon
- Font, Enrique & Molina-Borja, Miguel. (2004). Ultraviolet reflectance of color patches in Gallotia galloti from Tenerife, Canary Islands. https://www.researchgate.net/publication/270217243_Ultraviolet_reflectance_of_color_patches_in_Gallotia_galloti_from_Tenerife_Canary_Islands
- Prötzel, David & Heß, Martin & Scherz, Mark & Schwager, Martina & Van 't Padje, Anouk & Glaw, Frank. (2018). Widespread bone-based fluorescence in chameleons. Scientific Reports. 8. 10.1038/s41598-017-19070-7. https://www.researchgate.net/publication/322507066_Widespread_bone-based_fluorescence_in_chameleons
- Cox, Jonathan & Fitzpatrick, Benjamin. (2023). Biofluorescent sexual dimorphism revealed in a southern Appalachian endemic salamander, Plethodon metcalfi. Scientific Reports. 13. 10.1038/s41598-023-29051-8. https://www.researchgate.net/publication/368981642_Biofluorescent_sexual_dimorphism_revealed_in_a_southern_Appalachian_endemic_salamander_Plethodon_metcalfi
- Lamb, JY, Davis, MP Las salamandras y otros anfibios brillan con biofluorescencia. Sci Rep 10 , 2821 (2020). https://doi.org/10.1038/s41598-020-59528-9
- Thompson, Michelle & Saporito, Ralph & Ruiz-Valderrama, Diego & Medina Rangel, Guido & Donnelly, Maureen. (2019). A field-based survey of fluorescence in tropical tree frogs using an LED UV-B flashlight. https://www.researchgate.net/publication/336968084_A_field-based_survey_of_fluorescence_in_tropical_tree_frogs_using_an_LED_UV-B_flashlight
- Kohler, Allison & Olson, Erik & Martin, Jonathan & Anich, Paula. (2019). Ultraviolet fluorescence discovered in New World flying squirrels (Glaucomys). Journal of Mammalogy. https://www.researchgate.net/publication/330637392_Ultraviolet_fluorescence_discovered_in_New_World_flying_squirrels_Glaucomys
- Lim, Matthew & Land, M. & Li, Daiqin. (2007). Sex-Specific UV and Fluorescence Signals in Jumping Spiders. Science (New York, N.Y.). https://www.researchgate.net/publication/6547750_Sex-Specific_UV_and_Fluorescence_Signals_in_Jumping_Spiders
- Thomas, Daniel & Mcgoverin, Cushla & Mcgraw, Kevin & James, Helen & Madden, Odile. (2013). Vibrational spectroscopic analyses of unique yellow feather pigments (spheniscins) in penguins. https://www.researchgate.net/publication/236068133_Vibrational_spectroscopic_analyses_of_unique_yellow_feather_pigments_spheniscins_in_penguins
- Weidensaul, Scott & Colvin, Bruce & Brinker, David & Huy, J.. (2011). Use of Ultraviolet Light as an Aid in Age Classification of Owls. https://www.researchgate.net/publication/232672438_Use_of_Ultraviolet_Light_as_an_Aid_in_Age_Classification_of_Owls
- Camacho, Carlos & Negro, Juan & Redondo García, Iraida & Palacios Ojeda, Sebastián & Sáez-Gómez, Pedro. (2019). Correlates of individual variation in the porphyrin-based fluorescence of red-necked nightjars (Caprimulgus ruficollis). https://www.researchgate.net/publication/337923811_Correlates_of_individual_variation_in_the_porphyrin-based_fluorescence_of_red-necked_nightjars_Caprimulgus_ruficollis
- Hausmann, Franziska & Arnold, Kathryn & Marshall, N & Owens, Ian. (2003). Ultraviolet signals in birds are special. https://www.researchgate.net/publication/10896416_Ultraviolet_signals_in_birds_are_special
- Galván, Ismael & Camarero, Pablo & Mateo, Rafael & Negro, Juan. (2016). Porphyrins produce uniquely ephemeral animal colouration: A possible signal of virginity. https://www.researchgate.net/publication/310060626_Porphyrins_produce_uniquely_ephemeral_animal_colouration_A_possible_signal_of_virginity
- Wails, Christy & Gruber, Eva & Slattery, Ethan & Smith, Lucy & Major, Heather. (2017). Glowing in the light: Fluorescence of bill plates in the Crested Auklet (Aethia cristatella). https://www.researchgate.net/publication/301345953_Glowing_in_the_light_Fluorescence_of_bill_plates_in_the_Crested_Auklet_Aethia_cristatella
- Dunning, Jamie & Diamond, Tony & Christmas, Steve & Cole, Emma-Louise & Holberton, Rebecca & Jackson, Hannah & Kelly, Kevin & Brown, Dean & Rivera, Indira & Hanley, Daniel. (2019). Photoluminescence in the bill of the Atlantic Puffin Fratercula arctica. https://www.researchgate.net/publication/330676537_Photoluminescence_in_the_bill_of_the_Atlantic_Puffin_Fratercula_arctica
- Wilkinson, B.P., Johns, M.E. and Warzybok, P. (2019), Fluorescent ornamentation in the Rhinoceros Auklet Cerorhinca monocerata. https://onlinelibrary.wiley.com/doi/abs/10.1111/ibi.12715
- Boles, W. E. 1990 Glowing parrots—need for a study of hid-den colours. Birds Int. 3, 76–79.
- Pearn, S. M., Bennett, A. T. D. & Cuthill, I. C. 2001 Ultra-violet vision, fluorescence and mate choice in a parrot, the budgerigar Melopsittacus undulates. https://www.jstor.org/stable/3068090?seq=1
- Arnold, Kathryn & Owens, Ian & Marshall, N. (2002). Fluorescent Signaling in Parrots. https://www.researchgate.net/publication/11580061_Fluorescent_Signaling_in_Parrots/citations
- Evtukh, Grigory. (2019). Fluorescence among Fraterculinae subfamily. https://www.researchgate.net/publication/333658414_Fluorescence_among_Fraterculinae_subfamily
- Martin, Rene & Carr, Emily & Sparks, John. (2025). Does biofluorescence enhance visual signals in birds-of-paradise?. https://www.researchgate.net/publication/388921998_Does_biofluorescence_enhance_visual_signals_in_birds-of-paradise
- Ödeen, A., Håstad, O. & Alström, P. Evolution of ultraviolet vision in the largest avian radiation - the passerines (2011). https://doi.org/10.1186/1471-2148-11-313
- Marshall J, Johnsen S. 2017 Fluorescence as a means of colour signal enhancement https://royalsocietypublishing.org/doi/10.1098/rstb.2016.0335
- Nicolaï MPJ, Bok MJ, Abalos J, D’Alba L, Shawkey MD, Goldenberg J. 2024 The function and consequences of fluorescence in tetrapods. https://pubmed.ncbi.nlm.nih.gov/38814876/
- CUTHILL, I. C., J. C. PARTRIDGE, A. T. D. B ENNETT, S. C. CHURCH , N. S. H ART, AND S. H UNT. 2000. Ultraviolet vision in birds.
- https://www.sciencedirect.com/science/article/abs/pii/S0065345408601059?via%3Dihub
- Pierre Jouventin, Paul M. Nolan, Jonas Örnborg, F. Stephen Dobson, Ultraviolet Beak Spots in King and Emperor Penguins https://doi.org/10.1093/condor/107.1.144
- Jouventin P, Couchoux C, Dobson FS (2009) UV signals in penguins https://doi.org/10.1007/s00300-008-0564-3
- Dobson, Frederick & Nicolaus, Marion & Karels, Tim & Mcgraw, Kevin & Jouventin, Pierre. (2010). Mutual Mate Choice for Colorful Traits in King Penguins. https://www.researchgate.net/publication/229872073_Mutual_Mate_Choice_for_Colorful_Traits_in_King_Penguins
- Hadden, Peter & Vorobyev, Misha & Hadden, William & Bonadonna, Francesco & McGhee, Charles & Zhang, Jie. (2023). Can penguins (Spheniscidae) see in the ultraviolet spectrum? https://www.researchgate.net/publication/373235210_Can_penguins_Spheniscidae_see_in_the_ultraviolet_spectrum
- Longrich, Nicholas. (2010). The function of large eyes in Protoceratops: A nocturnal ceratopsian?. New Perspectives on Horned Dinosaurs, the Royal Tyrrell Museum Ceratopsian Symposium. 308-327.
- Witmer LM, Thomason JJ. 1995. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils.
- Twyman H, Valenzuela N, Literman R, Andersson S, Mundy NI. 2016. Seeing red to being red: conserved genetic mechanism for red cone oil droplets and co-option for red coloration in birds and turtles.
- Kelber, Almut & Vorobyev, Misha & Osorio, Daniel. (2003). Animal colour vision - Behavioural tests and physiological concepts. https://www.researchgate.net/publication/8064482_Animal_colour_vision__Behavioural_tests_and_physiological_concepts
- Chiari Y, Cahais V, Galtier N, Delsuc F. 2012. Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria). https://bmcbiol.biomedcentral.com/articles/10.1186/1741-7007-10-65
- Crawford NG, Parham JF, Sellas AB, Faircloth BC, Glenn TC, Papenfuss TJ, Henderson JB, Hansen MH, Simison WB. 2015. A phylogenomic analysis of turtles. https://tsjok45.wordpress.com/wp-content/uploads/2012/11/turtles-1-s2-0-s1055790314003819-main-1.pdf
- Hone D,W E, Naish D, Cuthill IC.2011. Doesmutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs?
- Pascal, Godefroit & Sinitsa, Sofia & Danielle, Dhouailly & Bolotsky, Yuri & Sizov, Alexander & McNamara, Maria & Benton, Michael & Spagna, Paolo. (2014). Dinosaur evolution. A Jurassic ornithischian dinosaur from Siberia with both feathers and scales. https://www.researchgate.net/publication/264247851_Dinosaur_evolution_A_Jurassic_ornithischian_dinosaur_from_Siberia_with_both_feathers_and_scales
- Yang, Zixiao & Jiang, Baoyu & McNamara, Maria & Kearns, Stuart & Pittman, Michael & Kaye, Thomas & Orr, Patrick & Xu, Xing & Benton, Michael. (2019). Pterosaur integumentary structures with complex feather-like branching. Nature Ecology & Evolution. https://www.researchgate.net/publication/329715712_Pterosaur_integumentary_structures_with_complex_feather-like_branching
No comments:
Post a Comment